Genetic diversity
Theory and practice
Per Unneberg
NBIS
15-Nov-2023
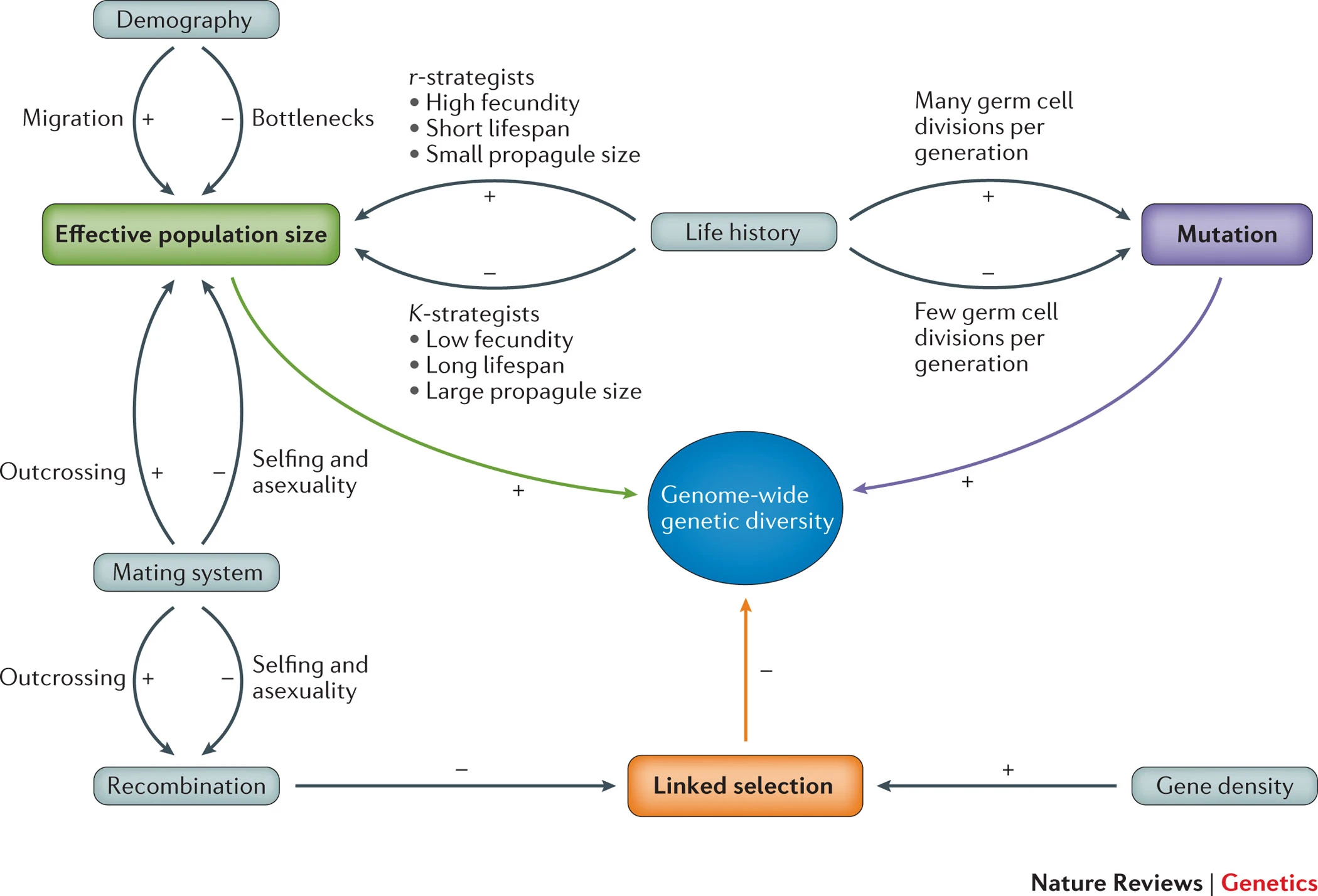
What determines diversity levels?

The usual questions:
What evolutionary forces maintain genetic diversity in natural populations? How do diversity levels relate to census population sizes…? Do low levels of diversity limit adaptation to selective pressures?
Leffler et al. (2012)
After allozyme era, the study of genetic diversity was largely neglected due to lack of genome-wide data, but with advent of population genomics becoming a hot topic again.
Ellegren & Galtier (2016)
Lewontin’s paradox: genetic diversity range smaller than variation among species in population size
Genetic diversity in conservation biology

…no global relationship between numerically coded IUCN extinction risk categories and estimated heterozygosity…
Low genetic diversity symptom of past genetic drift inbreeding (higher levels of homozygosity), caused by low \(N_e\)
García-Dorado & Caballero (2021)
However: if population decline is rapid, may be too little time for inbreeding to occur \(\Rightarrow\) genetic diversity within species not necessarily aligned to extinction risk
Lewis (2023)
Many programs treat missing data as invariant

Diversity:
\[ \pi = \frac{\sum_{i<j}k_{ij}}{n \choose 2} \]
Divergence:
\[ d_{XY} = \frac{1}{n_Xn_Y}\sum_{i=1}^{n_X}\sum_{j=1}^{n_Y}k_{ij} \]
Here, \(n\) is the number of samples, \(k_{ij}\) tally of allelic differences between two haplotypes within (\(\pi\)) a population or between (\(d_{XY}\)) populations
Missing data may bias diversity measures downwards

Nucleotide diversity landscapes

Genetic basis of adaptation and genome scans

Fundamental questions:
- How many genes are involved in the evolution of adaptive traits?
- What is the distribution of phenotypic effects among successive allelic substitutions?
- Is adaptation typically based on standing variation or new mutations?
- What is the relative importance of additive vs. nonadditive effects on adaptive trait variation?
- And what is the relative importance of structural vs. regulatory changes in phenotypic evolution?
Storz (2005), Fig 1
Example
vcftools --gzvcf allsites.vcf.gz --weir-fst-pop PUN-Y.txt \
--weir-fst-pop PUN-R.txt \
--fst-window-size 1000
csvtk plot line --tabs out.windowed.weir.fst \
-x BIN_START -y MEAN_FST \
--point-size 2 --xlab "Position (bp)" \
--ylab "Fst" --title "LG4: PUN-Y vs PUN-R" \
--width 9.0 --height 3.5 --scatter \
> out.windowed.weir.fst.mean.png
Z-scores can help identifying outliers

Raw data can be converted to Z-scores to highlight outliers. A Z-score is a measure of how far a data point is from the mean in terms of the number of standard deviations:
\[ Z = \frac{X - \mu}{\sigma} \]
Threshold of a couple of standard deviations common.
LD decay and choice of window size

Properties of genetic variation and inferred demographic history in sampled A. millepora. Fuller et al. (2020), Figure 2. Upper left plot illustrates LD as a function of physical distance. Here, choosing a window size 20-30kb would ensure that most windows are independent.
Dissecting differentiation landscapes

Monkeyflower genomic landscape
