
Selection
Selection
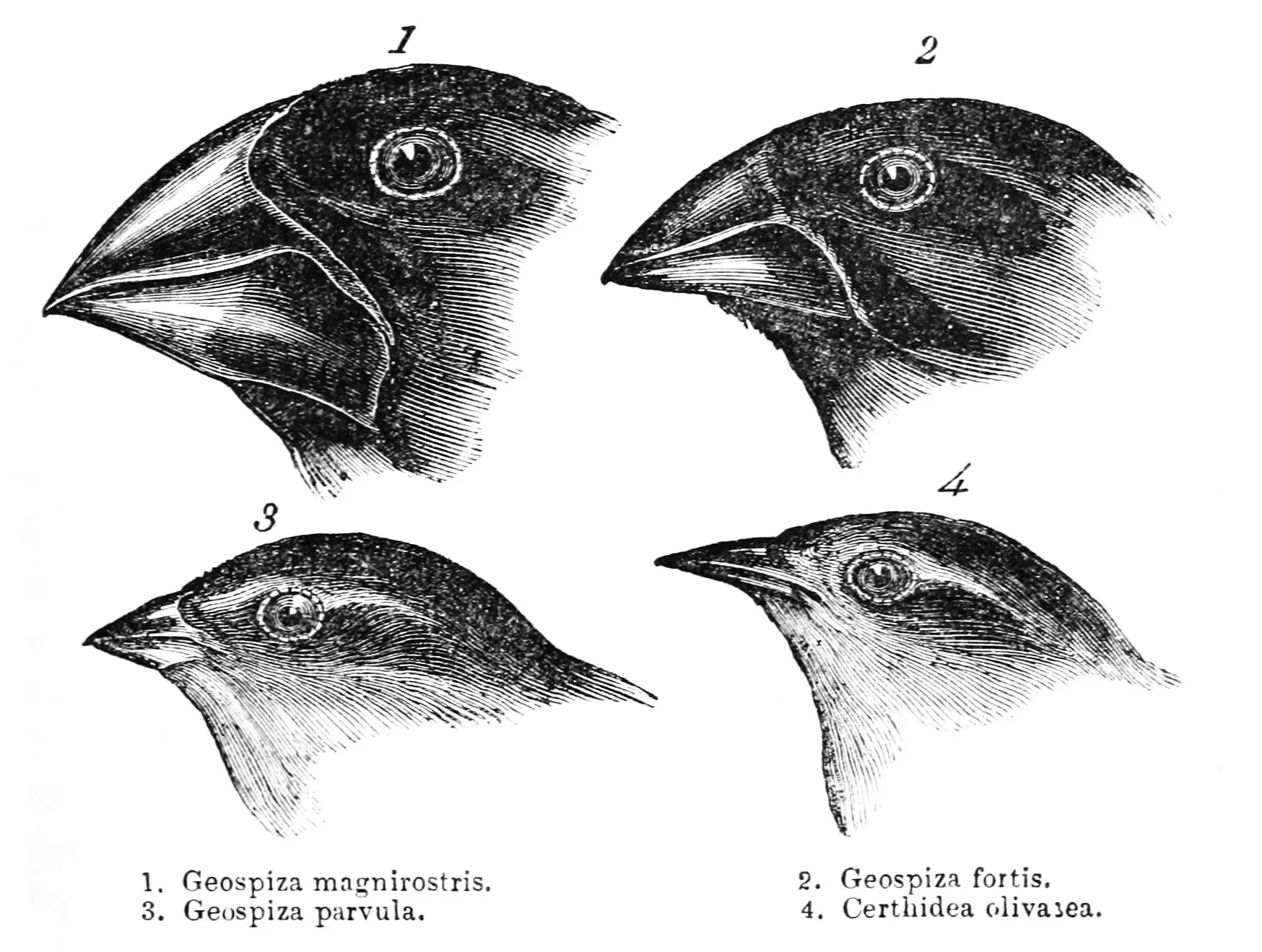
Selection and fitness
Much confusion exists in the literature regarding how various types of selection are defined, in particular because some of the terminology is used slightly differently within different scientific communities (Nielsen, 2005)
\begin{matrix} \mathrm{Genotype} & AA & Aa & aa \\ \mathrm{Frequency\ in\ newborns} & p^2 & 2pq & q^2\\ \mathrm{Viability} & w_{AA} & w_{Aa} & w_{aa}\\ \mathrm{Frequency\ after\ selection} & p^2w_{AA} / \bar{w} & 2pqw_{Aa} / \bar{w} & q^2w_{aa} / \bar{w} \\ \mathrm{Relative\ fitness} & 1 & 1-hs & 1-s\\ \end{matrix}
where \bar{w} = p^2w_{AA} + 2pqw_{Aa} + q^2w_{aa} is the mean fitness.
| h=0 | A dominant, a recessive |
| h=1 | a dominant, A recessive |
| 0<h<1 | incomplete dominance |
| h<0 | overdominance (heterozygote advantage) |
| h>1 | underdominance |
Notation follows Gillespie (2004), pp. 61–64.
The most important equation in population genetics
p^\prime - p = \Delta_sp = \frac{pq[p(w_{AA} - w_{Aa}) + q(w_{Aa} - w_{aa})]}{p^2w_{AA} + 2pqw_{Aa} + q^2w_{aa}}


Selection and drift - population size matters

In red region (|N_es|<0.05) the probability of fixation is within 10% of neutral fixation.
Consequence: for any population size there exists range of selection coefficients where mutant alleles \approx neutral (effective neutrality).
Direct selection can be inferred from protein substitutions
For genes, the ratio of nonsynonymous to synonymous substitutions can tell us about protein evolution:
Synonymous substitution
Protein L
DNA --- CTT ---
*
DNA --- CTC ---
Protein LNonsynonymous substitution
Protein L
DNA --- CTT ---
*
DNA --- CHT ---
Protein H- \mathbf{d_N/d_S << 1}
- negative (purifying) selection
- \mathbf{d_N/d_S < 1}
- majority nonsynonymous deleterious, some advantageous
- \mathbf{d_N/d_S = 1}
- neutral or mix neutral / advantageous / deleterious mutations
- \mathbf{d_N/d_S > 1}
- positive selection

Not all mutations fall in genes. Methods for detecting direct selection not applicable to studying selection on single mutation, or e.g., balancing. This requires looking for specific patterns of diversity surrounding locus under selection.
Linked selection reduces diversity at neighbour loci

Example of a selective sweep. If a sweep completes at a locus, it will become monomorphic, as will the neighbouring sites. Mutation could reintroduce variation. Recombination could increase diversity in neighbourhood, but in a manner that depends on the distance from the locus under selection.
The effect of a selective sweep on diversity
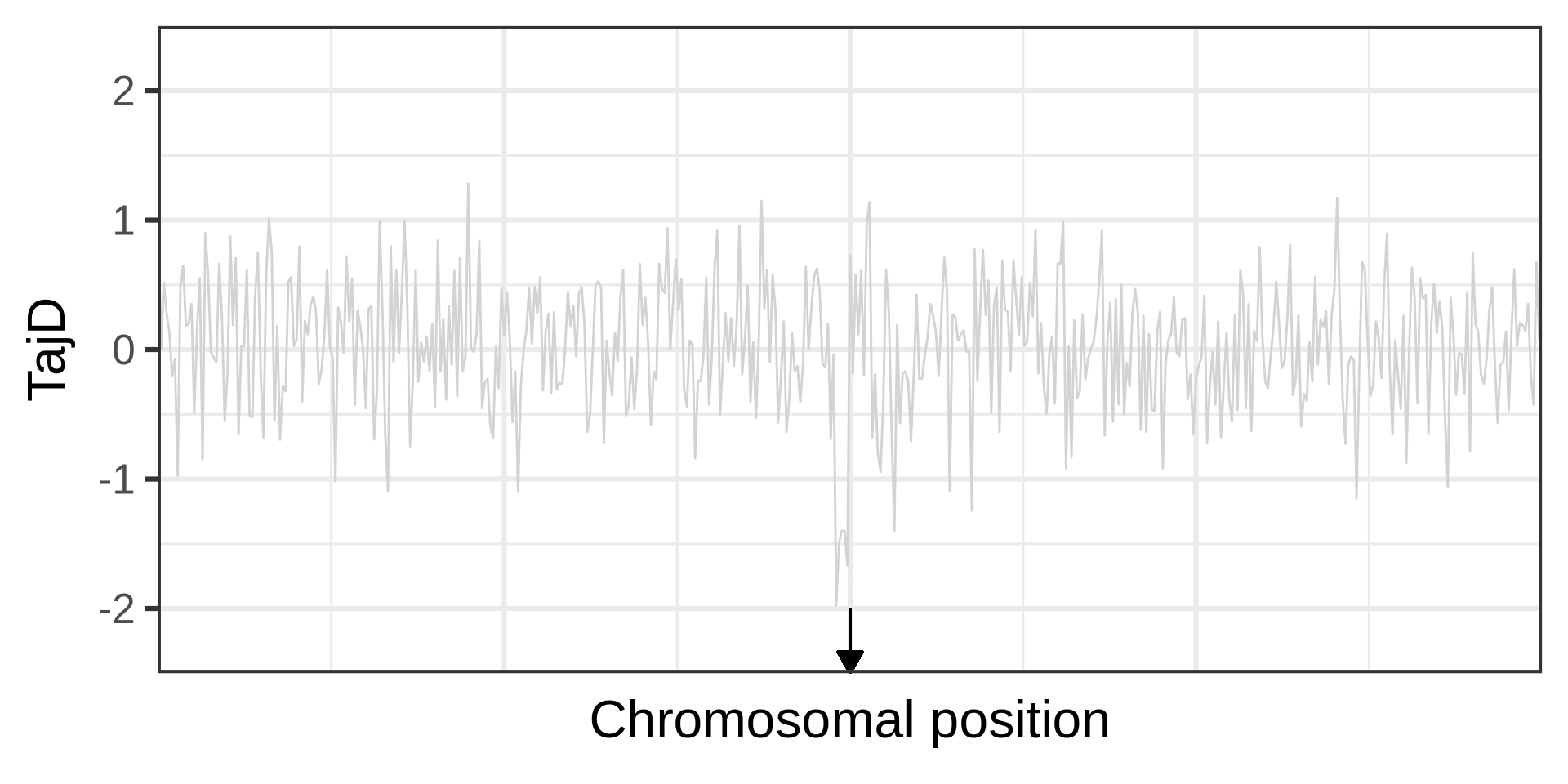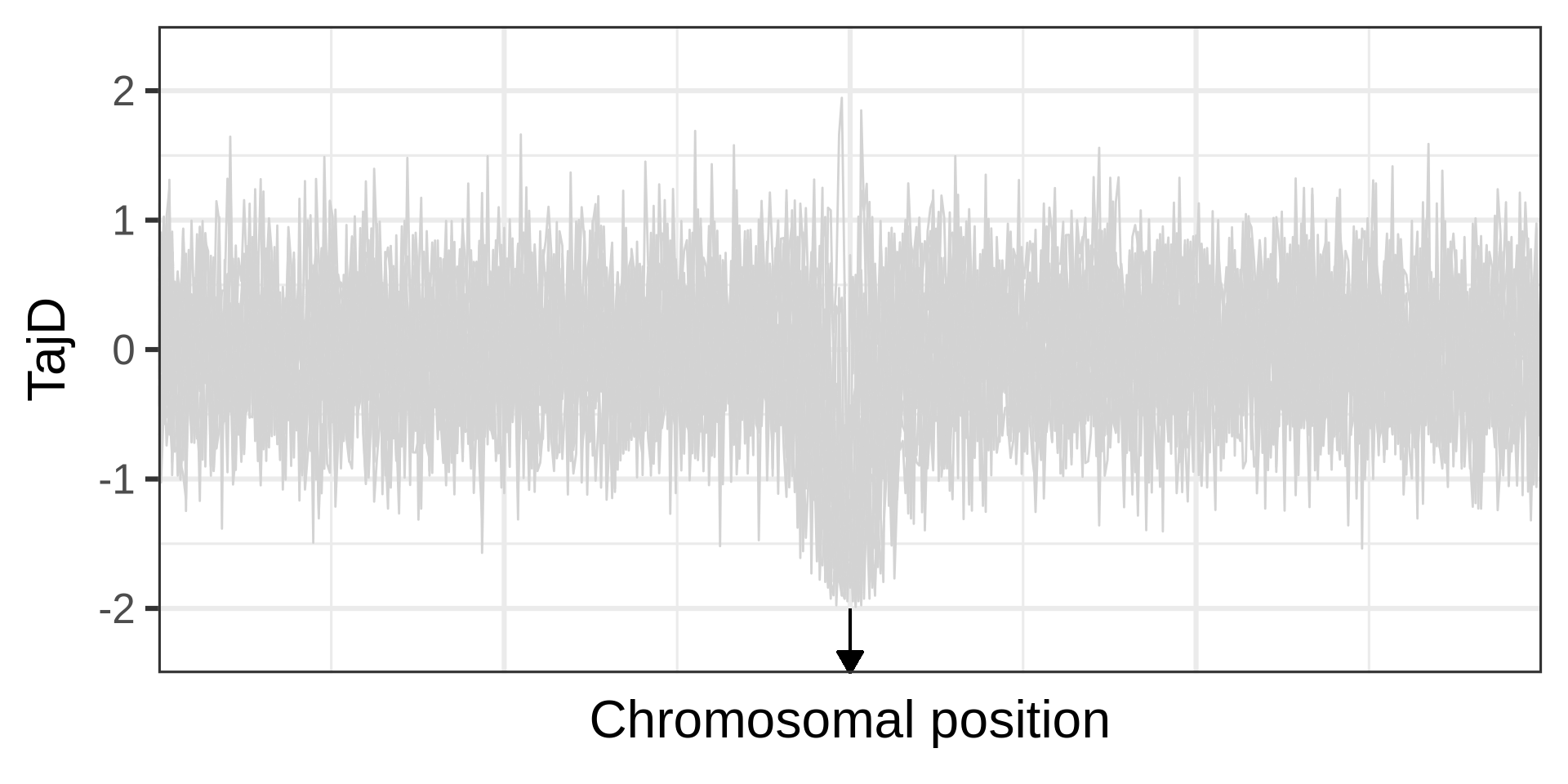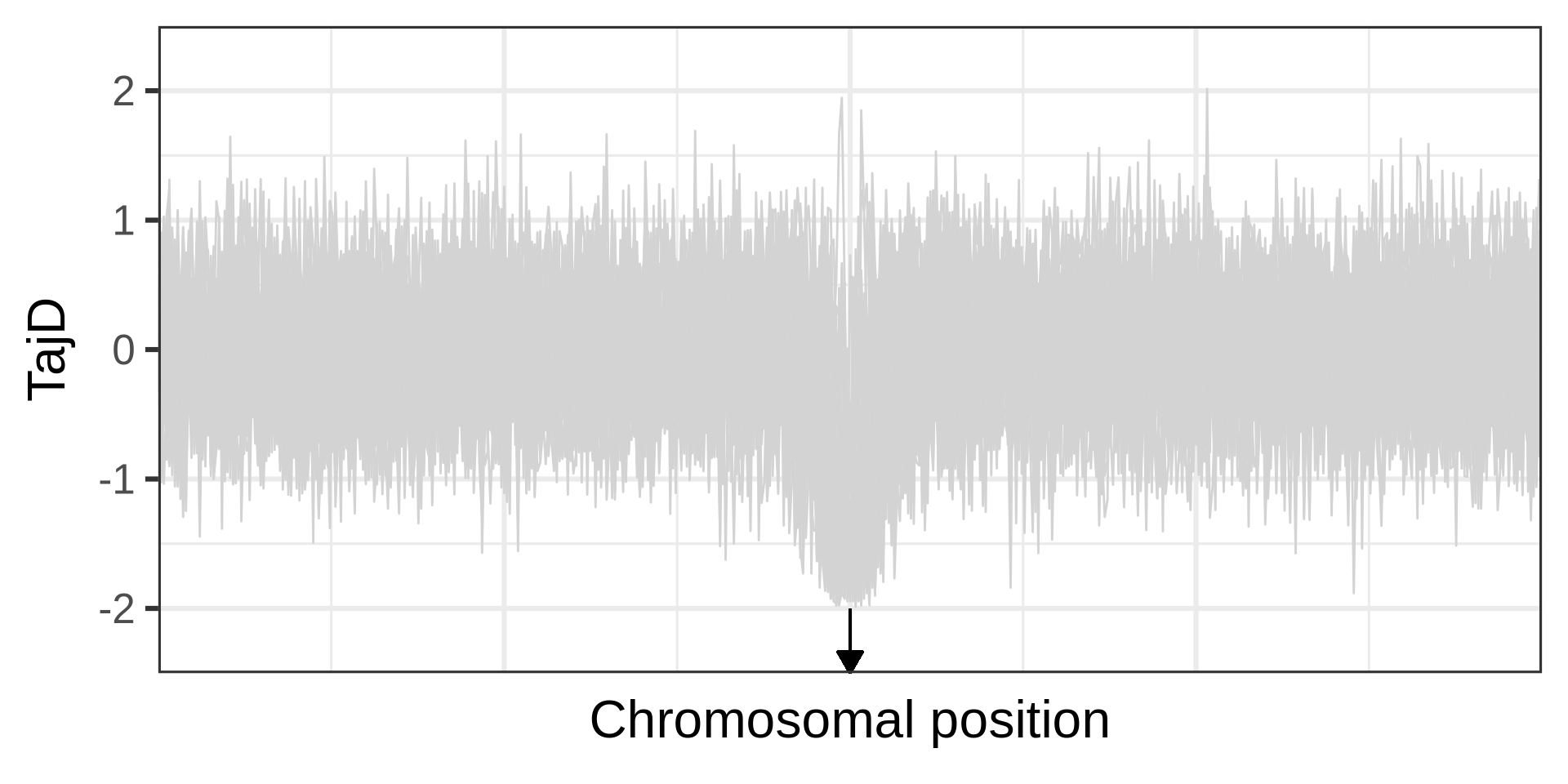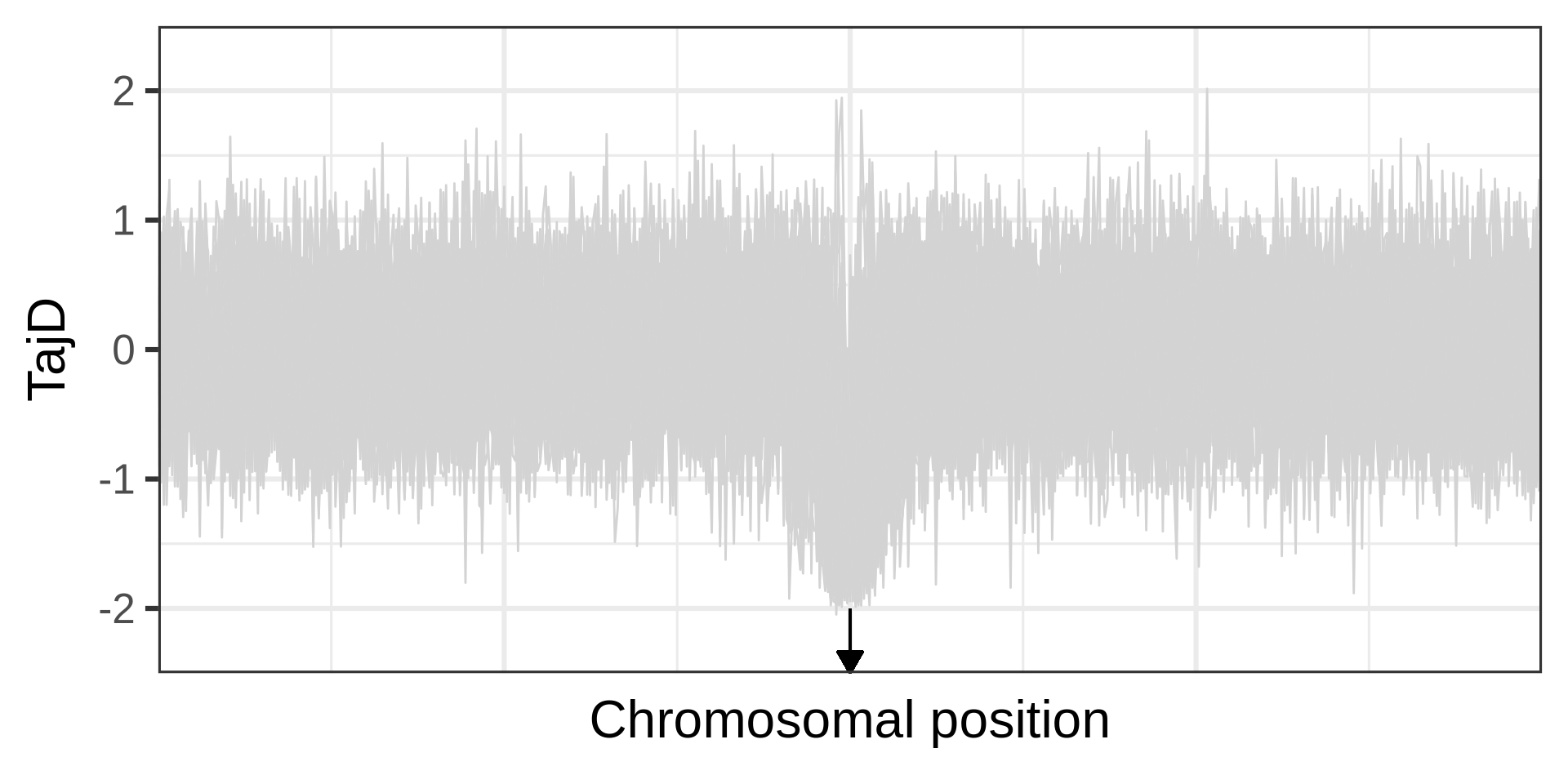
The effect of a selective sweep on diversity

The effect of a selective sweep on diversity

The phases of a selective sweep

Amount of diversity depends on fixation time. A neutral locus fixes in 4N_e generations; for s=0.0001, it takes approximately 0.29N_e generations.
Selections changes the genealogy (different topology, shorter branches), an aspect used in many linkage-based tests for selection.
Linked selection may constrain levels of diversity

Hitchhiking
- alleles linked to locus under selection “hitchhike” to high frequencies (Smith & Haigh, 1974)
- evidence: positive correlation between putative neutral diversity and recombination (Corbett-Detig et al., 2015)
Background selection
- loci linked to a deleterious locus will be purged from population and thus reduce diversity (Charlesworth et al., 1993)
- similar patterns to hitchhiking
Bibliography
Charlesworth, B., & Charlesworth, D. (2010). Elements of Evolutionary Genetics. Roberts and Company Publishers.
Charlesworth, B., Morgan, M. T., & Charlesworth, D. (1993). The Effect of Deleterious Mutations on Neutral Molecular Variation. Genetics, 134(4), 1289–1303.
Corbett-Detig, R. B., Hartl, D. L., & Sackton, T. B. (2015). Natural Selection Constrains Neutral Diversity across A Wide Range of Species. PLOS Biology, 13(4), e1002112. https://doi.org/10.1371/journal.pbio.1002112
Gillespie, J. H. (2004). Population Genetics: A Concise Guide (2nd edition). Johns Hopkins University Press.
Hahn, M. (2019). Molecular Population Genetics (First). Oxford University Press.
Lynch, M. (2007). The origins of genome architecture. Sinauer Associates.
Nielsen, R. (2005). Molecular Signatures of Natural Selection. Annual Review of Genetics, 39(1), 197–218. https://doi.org/10.1146/annurev.genet.39.073003.112420
Smith, J. M., & Haigh, J. (1974). The hitch-hiking effect of a favourable gene. Genetics Research, 23(1), 23–35. https://doi.org/10.1017/S0016672300014634